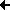Chloroplasten enthalten auch genetische Information. Ihre Bedeutung als Erbträger geht auf frühere Arbeiten von E. Baur (1909) und C. Correns (1909) zurück. Die molekulargenetische Phase der Forschung setzte 1962 mit dem definitiven Nachweis von DNS in Chloroplasten ein (H. Ris und W. Plaut, University of Wisconsin, Madison).
Für diese Gene würde die Chloroplasten-DNS rein rechnerisch gerade noch ausreichen, da sie bei fast allen höheren Pflanzen etwa 150.000 Basenpaare enthält. Doch in Chloroplasten laufen noch weitere Aktivitäten ab, deshalb würden weiterhin noch Enzyme für die folgenden Prozesse benötigt:
Wechselwirkung zwischen plastiden- und kern-codierten Proteinen
Schon allein dadurch wird deutlich, dass viele Enzyme aus dem Cytosol in die Chloroplasten importiert werden müssen. Es besteht also ein Gleichgewicht zwischen chloroplasten-codierten und kern-codierten Funktionen. Die Chloroplasten-DNS wird durch kern-codierte Enzyme repliziert. Diese werden im Cytosol gebildet und anschließend in die Chloroplasten überführt. Die Chloroplastenribosomen enthalten rRNS, die chloroplasten-codiert ist, und Proteine, die einerseits chloroplasten-codiert, andererseits kern-codiert sind.
Durch einen Versuch mit spezifischen  Inhibitoren (Antibiotika) lässt sich entscheiden, ob ein Protein im Cytosol oder in den Chloroplasten gebildet wird: Streptomycin und Chloramphenicol hemmen die Proteinbiosynthese an Chloroplastenribosomen. Cyoloheximid dagegen hemmt die Proteinbiosynthese an den Ribosomen des Cytosols. Setzt man diese Hemmstoffe ein, kann man die Herkunft und damit den Syntheseort einer Reihe von Proteinen ermitteln.
Inhibitoren (Antibiotika) lässt sich entscheiden, ob ein Protein im Cytosol oder in den Chloroplasten gebildet wird: Streptomycin und Chloramphenicol hemmen die Proteinbiosynthese an Chloroplastenribosomen. Cyoloheximid dagegen hemmt die Proteinbiosynthese an den Ribosomen des Cytosols. Setzt man diese Hemmstoffe ein, kann man die Herkunft und damit den Syntheseort einer Reihe von Proteinen ermitteln.
Darüber hinaus kann man diese Frage durch den Einsatz von Mutanten klären: durch genetische Analyse kann entschieden werden, ob ein Defekt dem Chloroplasten- oder dem Kerngenom zuzuschreiben ist, und durch eine biochemische Analyse (Auftrennung der Proteine, beispielsweise durch Gel-Elektrophorese) kann er auf Veränderung eines bestimmten Proteins zurückgeführt werden.
Eine dritte Möglichkeit wäre die Analyse der Proteinbiosynthese in isolierten Chloroplasten oder in invitro-Proteinbiosynthese-Systemen, die aus Bestandteilen der Chloroplasten aufgebaut werden können.
Ein wichtiger Aspekt ist der Befund, dass die oligomeren Proteine der  Photosynthesemembran aus Untereinheiten bestehen, die teilweise chloroplasten-codiert, zum Teil aber auch kern-codiert sind. Sie sind also gute Beispiele, um die Wechselwirkung zwischen den beiden Genomen zu untersuchen.
Photosynthesemembran aus Untereinheiten bestehen, die teilweise chloroplasten-codiert, zum Teil aber auch kern-codiert sind. Sie sind also gute Beispiele, um die Wechselwirkung zwischen den beiden Genomen zu untersuchen.
Ungeklärt bleibt dabei jedoch die Frage, wie sich eine solche Kooperation im Verlauf der Evolution herausgebildet hat. Gehen wir von der Endosymbiontenhypothese und einigen vergleichbaren Befunden der Molekularbiologie ("wandernde" / "springende" Gene) aus, ist es durchaus plausibel anzunehmen, dass ursprünglich die gesamte benötigte genetische Information in der Chloroplasten-DNS kodiert war, dass dann aber Teile aus den Chloroplasten in den Kern wanderten und seither als Kerngene repliziert und exprimiert werden.
An dieser Grafik kann man verdeutlichen, welche Proteine im Chloroplasten kern- bzw. chloroplasten-codiert sind. TS ist die Transitsequenz, der Abschnitt der Polypeptidkette, der für das Durchschleusen des Moleküls durch die Membran benötigt und anschließend abgespalten wird.
Transportmechanismen durch die Chloroplastenhülle
Ein Protein, das im Cytosol gebildet wird, muss mindestens zwei Membranen durchdringen: die äußere und die innere Plastidenmembran. Einige wenige Proteine, die kern-codiert sind, aber an der Innenseite der Thylakoiden aktiv sind, müssen sogar drei Membranen passieren. Man findet - zumindest bei höheren Pflanzen - auch keine Ribosomen an der äußeren Chloroplastenmembran, die Proteinbiosynthese durch die Membran hindurch kann also nicht zutreffen. Folglich muss ein Transport der gebildeten Proteine existieren.
B. Dibberstein, G. Blobel und N.-H. Chua (Rockefeller University, 1977) entdeckten, dass Proteine, die im Cytosol gebildet werden, um einiges größer sind als die später funktionellen Formen in den Chloroplasten.
Den zusätzlichen Abschnitt belegte man mit dem Begriff Transitsequenz bzw. Transfersequenz. Diese muss offensichtlich Rezeptoren an der Außenseite der Chloroplasten erkennen. Ferner muss es spezifische Carrier geben, um Proteine zu binden und durch die Membran zu translokieren. Letztlich wird eine Endoprotease benötigt, um die Transportform des Proteins in die aktive Form zu bringen.
Eine Vorbehandlung von Chloroplasten mit einer Protease unterbindet jegliche Proteinaufnahme: die benötigten Rezeptoren für die die Transitsequenz und die Carrier wurden außer Kraft gesetzt. Ohne sie kann kein Protein eingeschleust werden.
Dieser Mechanismus erklärt jedoch nur, wie die äußere Chloroplastenmembran überwunden wird. R. Douce entdeckte 1973, dass die Abstände der beiden Membranen der Chloroplastenhülle nicht konstant sind. Die beiden Membranen schwingen, so dass eine oszillierende Bewegung entsteht. In regelmäßigen Zeitabständen berühren sie sich kurzfristig punktuell und fusionieren reversibel. An solchen Stellen hätte ein Protein nur eine einzige Membran zu durchdringen.