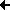Es stand nie in Frage, dass jeder der biochemischen Reaktionsschritte bei der Photosynthese durch je ein spezifisches Enzym katalysiert wird, doch es dauerte erstaunlich lange, bevor man erkannte, dass das  Chlorophyll und die anderen Pigmente proteingebunden sein müssen, also nur als Protein-Pigment-Komplex aktiv sein können. Die chromatographisch aufgetrennten und damit isolierten Pigmente sind für sich allein genommen nicht funktionstüchtig.
Chlorophyll und die anderen Pigmente proteingebunden sein müssen, also nur als Protein-Pigment-Komplex aktiv sein können. Die chromatographisch aufgetrennten und damit isolierten Pigmente sind für sich allein genommen nicht funktionstüchtig.
Die Pigment-Protein-Komplexe, die Proteine der Elektronentransportketten sowie die ATP-Synthetase, der Katalysator für die Synthese des ATP, sind integrale Bestandteile der Thylakoidmembranen, also der Photosynthesemembran. Die Positionen in den Membranen - zum Beispiel außen oder innen - und die relative Anordnung der Proteine zueinander sind die entscheidenden Voraussetzungen für Energieumwandlungen in der Photosynthese.
Benötigt für die Energieumwandlung werden völlig intakte, protonenundurchlässige Membranen, die  Kompartimente umschließen, so dass ein elektrochemischer Gradient zwischen innen und außen aufrechterhalten werden kann.
Kompartimente umschließen, so dass ein elektrochemischer Gradient zwischen innen und außen aufrechterhalten werden kann.
Proteine der Photosynthesemembran
Das Studium dieser Proteine setzte erst sehr spät ein, da es sich um membrangebundene Proteine handelt, deren Isolierung und Charakterisierung mit den klassischen Methoden der Proteinanalyse nicht gelang. Erst durch die Entwicklung empfindlicher Verfahren, wie der Gel-Elektrophorese, wurde es möglich, die Proteine aufzutrennen und als Banden in einem Gel zu identifizieren.
Ein zweiter Ansatz ist die Verwendung spezifischer Sonden, beispielsweise fluoreszenzmarkierter Antikörper. Mit ihrer Hilfe ist es möglich, zu entscheiden, ob ein bestimmtes Protein an der Innen- oder der Außenseite einer Membran angebracht ist. Sie erlauben weiterhin, das betreffende Protein aus einem Gemisch selektiv auszufällen, weil nur dieses den hochspezifischen Antigen-Antikörper-Komplex ausbilden kann.
Ein dritter Ansatz zur Charakterisierung der Photosynthesemembran ist die Analyse bestimmter Mutanten. Man kennt eine Reihe von Mutanten mit Photosynthesedefekten, die sich vier Klassen zuordnen lassen:
- Mutanten mit einem Defekt im Photosystem I
- Mutanten mit einem Defekt im Photosystem II
- Mutanten mit einem Defekt in der Photophosphorylierung
- Mutanten mit einem Defekt im Lichtsammlerkomplex
Fast alle Mutanten sind nicht durch den Verlust oder die Veränderung einer bestimmten Polypeptidkette gekennzeichnet, sondern gleich durch den Ausfall eines ganzen Komplexes: Durch Veränderung einer einzigen Polypeptidkette funktioniert die Zusammenlagerung der übrigen Polypeptidketten zu funktionalen Einheiten nicht mehr. Daran zeigt sich, wie eng die Wechselwirkung zwischen den einzelnen Polypeptidketten tatsächlich ist.
Als letztes Verfahren bietet sich noch die Elektronenmikroskopie an, die meist in Kombination mit der Gefrierätztechnik genutzt wird.
Die Sequenzierung von Membranproteinen bleibt jedoch nach wie vor schwierig. Nur über den Umweg der Nukleotidsequenzierung der entsprechenden Gene konnten in den letzten Jahren die Aminosäuresequenzen der meisten beteiligten Proteine ermittelt werden.
Chlorophyllbindende Proteine
Es wurden verschiedene chlorophyllbindende Proteine aus Photosynthesemembranen mehrerer Arten isoliert und charakterisiert. Die bekanntesten Proteine sind das P700-Chlorophyll-a-Protein 1 und das Lichtsammler-Chlorophyll-a/b-Protein 2. Beide sind stark hydrophobe integrale Membranproteine. Beide binden Chlorphyll a, doch nur das zweite darüber hinaus auch Chlorophyll b.
Das P700-Chlorophyll-a-Protein 1 enthält das Reaktionszentrum (P700) für das Photosystem I. Das heißt, eines der Chlorophyllmoleküle ist in einer spezifischen Konfiguration gebunden und befindet sich in einer Umgebung, durch die es sich von allen übrigen auch an dieses Protein gebundenen Chlorophyllmolekülen unterscheidet. Diese strukturelle Besonderheit bildet die Voraussetzung für die lichtbetriebene Anregung und damit die Induktion eines Elektronenflusses.
Das Lichtsammlerprotein ist ebenfalls häufig und weit verbreitet. Es wird vorwiegend mit dem Photosystem II assoziiert. Chlorophyll a und b werden in äquimolaren Mengen neben Lutein und beta-Carotin gebunden. Das Chlorophyll-Carotinoid-Verhältnis beträgt dabei etwa 3:1 bis 7:1.
Zu den Aufgaben des Proteins gehört die Lichtfallenfunktion (Antennenfunktion), also die Weiterleitung von Photonen an das Reaktionszentrum (P680).
ATP-Synthetase
Zur Synthese von ATP benötigt man eine ATP-Synthetase, auch ATP-Synthase genannt, die als Katalysator dient. Die ATP-Bildung beruht auf einem gleichartigen Mechanismus wie bei der Atmungskette.
E. Racker (Cornell University) konnte aus Thylakoidmembranen ein Enzym isolieren, das weitgehend der ATP-Synthetase aus Mitochondrien ähnelt. Unter dem Elektronenmikroskop ist es als ein gestielter Knopf zu erkennen. Der Knopf erhielt die Bezeichnung CF1, der Stiel, der zur Verankerung in der Membran dient, die Bezeichnung CF0.
Im Gegensatz zur ATP-Synthetase bei Mitochondrien sitzt der Komplex an der Außenseite der Thylakoidmembran. Die Phosphorylierung von ADP gelingt nur, wenn die ATP-Synthetase Bestandteil intakter, protonendurchlässiger Membranen ist. Wichtig ist dabei auch, dass die Membran zwei Kompartimente (Vesikelinneres und Umgebung) voneinander trennt.
Chemiosmotische Hyptothese
Die ATP-Synthetase besitzt die Fähigkeit, Energie, die bei einem Elektronenfluss frei wird, für die Bildung von ATP zu nutzen. P. Mitchell (Glynn Research Laboratories, England) stellte 1961 die Hypothese auf, dass die Energie, die beim Elektronentransport anfällt, in einem Protonengradienten gespeichert wird. Die Energie wäre damit nicht in Form chemischer Bindungen, sondern in Form elektrochemischer Gradienten konserviert.
Bewiesen wurde diese Hypothese 1966 von A. T. Jagendorf (Cornell University, Ithaca N.Y.): er isolierte Thylakoide und inkubierte sie solange in einem pH-Puffer, bis sich auf beiden Seiten der Membran der gleiche pH-Wert (4) eingestellt hatte. Danach überführte er die Thylakoide schlagartig in ein Medium mit dem pH-Wert 8, das zusätzlich ADP und Phosphat enthielt. Unmittelbar nach der Überführung kam es unter ATP-Bildung zu einem pH-Ausgleich zwischen innen und außen.
Damit wurde gezeigt, dass ein Protonenfluss durch die Membran hindurch zur ATP-Bildung genutzt wird. Dieser Mechanismus funktioniert nur dann, wenn
- die Membranen unversehrt sind
- alle Enzyme gleichgerichtet sind, so dass Protonen nur in eine Richtung befördert werden
Um zu verstehen, wie der Protonentransport durch die Membran funktioniert, ist es nützlich, sich die Formalitäten der ATP-Bildung näher anzusehen. ADP und Phosphat werden als Ausgangsstoffe benötigt. Die beiden Moleküle werden an getrennten, aber benachbarten Bindungsstellen des Enzymkomplexes angelagert. Um zu ATP zu gelangen, müssen ein H+ vom ADP und ein OH- vom Phosphat abgespalten werden.
Die beiden Ionen entfernen sich jedoch in entgegengesetzte Richtungen und verbinden sich, sobald sie den Komplex verlassen haben, mit den jeweiligen Gegenionen zu Wasser. Als Bilanz erhalten wir aber eine gerichtete Protonenwanderung. Dabei wandert aber nicht etwa ein Proton via ATP-Synthetase durch die Membran hindurch, sondern es wird auf der einen Seite ein neu gebildetes (abgespaltenes) Proton abgegeben, während auf der Gegenseite ein anderes Proton (durch OH-) neutralisiert wird.